Google vertaling, originele artikel link en pdf
https://www.nature.com/articles/s41588-023-01410-1
30 mei 2023
Abstract
Spontane coronaire arteriedissectie (SCAD) is een onderbelichte oorzaak van een hartinfarct dat voornamelijk vrouwen treft. Het is niet bekend in hoeverre SCAD genetisch verschilt van andere hart- en vaatziekten, waaronder atherosclerotische coronaire hartziekte (CAD). Hier presenteren we een genoombrede associatie-meta-analyse (1.917 gevallen en 9.292 controles) die 16 risicoloci voor SCAD identificeren. Integratieve functionele annotaties gaven prioriteit aan genen die waarschijnlijk worden gereguleerd in vasculaire gladde spiercellen en arteriële fibroblasten en die betrokken zijn bij extracellulaire matrixbiologie. Eén locus met het tissue factor-gen F3, dat betrokken is bij de initiatie van de bloedstollingscascade, lijkt specifiek te zijn voor SCAD-risico. Verschillende geassocieerde varianten hebben diametraal tegenovergestelde associaties met CAD, wat suggereert dat gedeelde biologische processen bijdragen aan beide ziekten, maar via verschillende mechanismen. We concluderen ook een oorzakelijke rol voor hoge bloeddruk bij SCAD. Onze bevindingen bieden nieuwe pathofysiologische inzichten met betrekking tot arteriële integriteit en weefselgemedieerde coagulatie in SCAD en vormen de basis voor toekomstige specifieke therapieën en preventies.
Voornaamst
Hart- en vaatziekten zijn de belangrijkste doodsoorzaak bij vrouwen, maar seksespecifieke aspecten van het risico op hartaandoeningen en acuut myocardinfarct (AMI) blijven onderbelicht1. Spontane coronaire arteriedissectie (SCAD) en atherosclerotische coronaire hartziekte (CAD) zijn beide oorzaken van acute coronaire syndromen die leiden tot AMI2,3,4,5,6. In tegenstelling tot CAD treft SCAD echter een jongere, overwegend vrouwelijke populatie7 en ontstaat het door de ontwikkeling van een hematoom, wat leidt tot dissectie van de coronaire tunica media met de uiteindelijke vorming van een vals lumen, in plaats van atherosclerotische plaque-erosie of -ruptuur8. SCAD is klinisch in verband gebracht met migraine9 en extra-coronaire arteriopathieën, waaronder fibromusculaire dysplasie (FMD)10,11,12,13. Co-existente coronaire atherosclerose komt echter niet vaak voor8,14. Hoewel de genetische basis van CAD steeds beter ingeburgerd raakt15, blijft de pathofysiologie van SCAD slecht begrepen4. De zoektocht naar zeer penetrante mutaties in kandidaatroutes of door sequencing heeft een lage opbrengst opgeleverd, vaak wijzend op genen die betrokken zijn bij andere klinisch niet-gediagnosticeerde erfelijke syndromen die zich manifesteren als SCAD16. Eerdere onderzoeken naar de impact van algemene genetische variatie op het risico op SCAD hebben vijf bevestigde risicoloci beschreven17,18,19,20.
In dit artikel hebben we een meta-analyse uitgevoerd van genoombrede associatiestudies (GWAS’s) bestaande uit 1.917 SCAD-gevallen en 9.292 controles van Europese afkomst. We identificeerden 16 risico-loci, waaronder 11 nieuwe associatiesignalen, die een substantiële polygene erfelijkheid voor deze ziekte aantonen. Belangrijk is dat we laten zien dat verschillende gemeenschappelijke genetische risico-loci voor SCAD worden gedeeld met CAD, maar een tegengesteld effect hebben en een andere genetische bijdrage van gevestigde cardiovasculaire risicofactoren. Deze bevindingen impliceren arteriële integriteit gerelateerd aan extracellulaire matrixbiologie, vasculaire tonus en weefselcoagulatie in de pathofysiologie van SCAD.
Resultaten
GWAS-meta-analyse en erfelijkheidsgraad van single-nucleotide polymorfisme
We hebben een GWAS-meta-analyse uitgevoerd van acht onafhankelijke case-control-onderzoeken (aanvullende figuren 1 en 2 en aanvullende tabel 1). Zestien loci vertoonden genoombrede significante signalen van associatie met SCAD, waarvan er 11 nieuw werden beschreven voor deze ziekte (tabel 1, figuur 1a, aanvullende tabel 2 en aanvullende figuur 3). Eén locus op chromosoom 4 (AFAP1) werd onlangs gemeld voor SCAD in de context van zwangerschap19 en er is nu bevestigd dat deze in het algemeen betrokken is bij SCAD (Tabel 1). De geschatte odds ratio’s van geassocieerde loci varieerden van 1,25 (95% betrouwbaarheidsinterval (BI) = 1,16–1,35) in ZNF827 op chromosoom 4 tot 2,04 (95% BI = 1,77–2,35) op chromosoom 21 nabij KCNE2 (Tabel 1). We rapporteren bewijs voor substantiële polygeniciteit voor SCAD met een geschatte op single-nucleotide polymorphism (SNP) gebaseerde erfelijkheidsgraad van meer dan 0,70 (h2SNP = 0,71 ± 0,11 op de aansprakelijkheidsschaal met behulp van linkage onevenwichtigheidsscore-regressie21 en h2SNP = 0,70 ± 0,12 met behulp van SumHer22; aanvullende tabel 3 ). De ECM1/ADAMTSL4-locus op chromosoom 1 was verantwoordelijk voor het grootste deel van de erfelijkheidsgraad voor SCAD in onze dataset (h2 = 0,028), gevolgd door de COL4A1/COL4A2-locus, die twee onafhankelijke GWAS-signalen bevatte (h2 = 0,022; aanvullende tabel 4 en aanvullende afb. 4). Over het algemeen schatten we dat de 16 loci ~ 24% van de totale op SNP gebaseerde erfelijkheidsgraad van SCAD verklaren (aanvullende tabel 4).
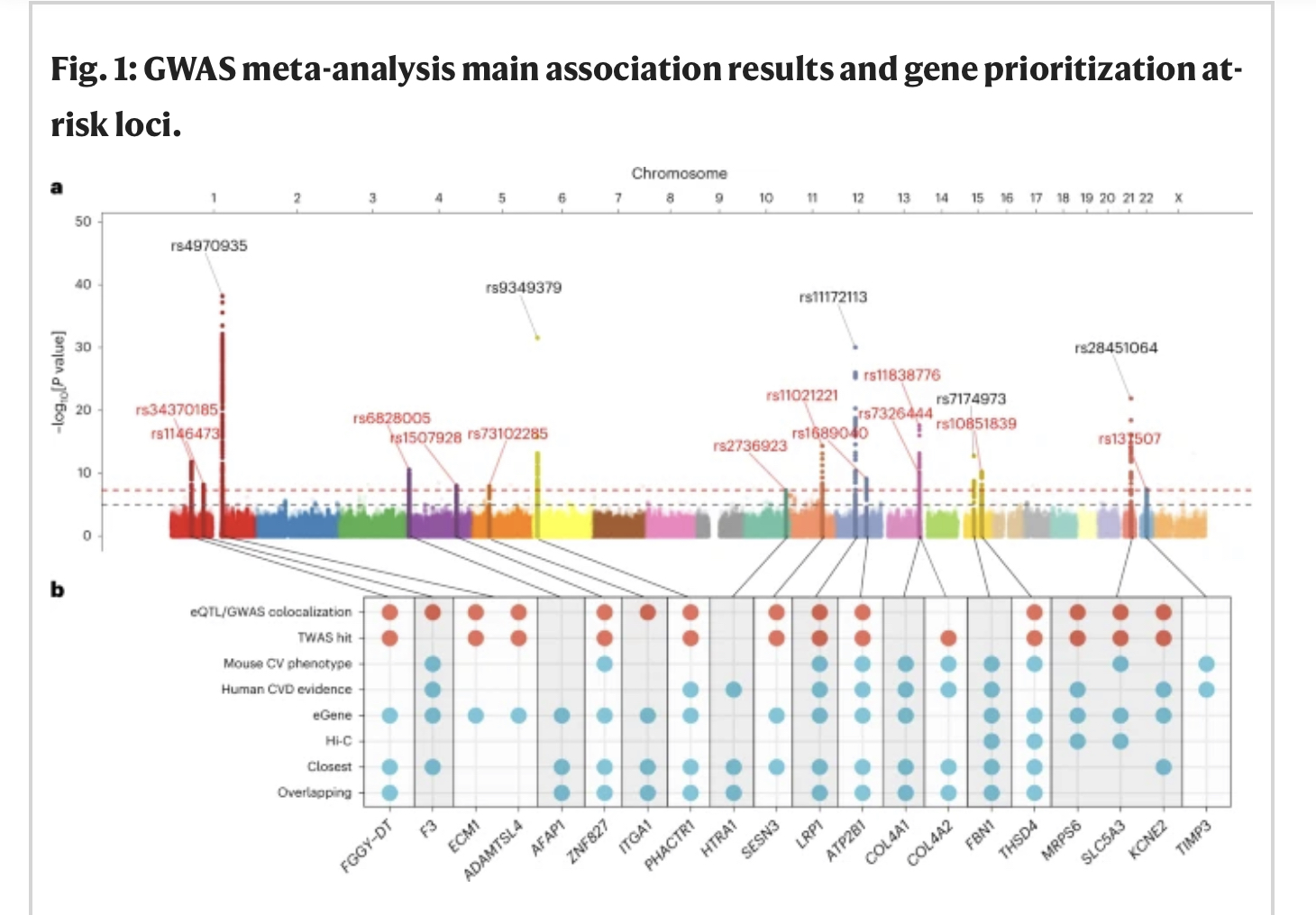
a, Manhattan-plotrepresentatie van op SNP gebaseerde associatie-meta-analyse in SCAD. De x as toont de genomische coördinaten en de y as toont de −log10[P waarde] verkregen door tweezijdige Wald-test. SNP’s rond genoombrede significante signalen (± 500 kb) zijn gemarkeerd. De labels tonen de rsID’s voor de leidende SNP’s, met nieuw geïdentificeerde loci in rood en voorheen bekende loci in zwart. De gestippelde rode lijn vertegenwoordigt genoombrede significantie (P = 5 × 10−8) en de grijze lijn suggestieve associatie (P = 10−5). b, Samenvatting van de strategie voor de annotatie van genprioritering. De stippen geven genen aan die voldoen aan een van de volgende acht criteria: (1) colokalisatie van SCAD-associatiesignaal en eQTL-associatie in de aorta, kransslagader, scheenbeenslagader, fibroblasten of volbloedmonsters (versie GTEx versie 8); (2) een TWAS-hit in een van de bovengenoemde weefsels; (3) een cardiovasculair (CV) fenotype in de gen-knockout-muis; (4) bestaand bewijs van genfunctie in pathofysiologie van hart- en vaatziekten (HVZ) bij mensen; (5) het gen is een eGene voor een nabij gelegen leidende SNP in de bovengenoemde GTEx-weefsels; (6) Hi-C-bewijs25 voor een promotor van het gen in een chromatinelus van menselijk aortaweefsel dat varianten omvat van de geloofwaardige reeks causale varianten; (7) het dichtstbijzijnde gen stroomopwaarts of stroomafwaarts van de leidende SNP; of (8) varianten in de geloofwaardige set van causale varianten in kaart in het gen. Criteria 1 en 2 (blauwe stippen) kregen een tienvoudige gewogen score ten opzichte van criteria 3–8. Genen met de meeste criteria kregen prioriteit in elke locus en worden hier getoond.
Functionele annotatie van varianten in SCAD-loci
We ontdekten dat SCAD-geassocieerde varianten significant verrijkt waren in versterkerkenmerken die specifiek zijn voor genexpressie in arteriële weefsels van ENCODE23 (bijvoorbeeld de aorta, tibiale slagader, thoracale aorta en kransslagader), evenals verschillende weefsels die rijk zijn aan gladde spiercellen ( bijvoorbeeld de dikke darm, dunne darm en baarmoeder) (aanvullende figuur 5). Op basis van recent gepubliceerde analyses van eencellig open chromatine in 30 volwassen weefsels , hebben we vastgesteld dat vasculaire gladde spiercellen (VSMC’s) en fibroblasten de meest verrijkte celtypen waren voor SCAD-geassocieerde loci tussen clusters die worden weergegeven in aorta- en tibiale arterie-datasets (Fig. 2a en aanvullende figuur 6). Consequent bevatten alle SCAD-locus op één na ten minste één variant die overlapte met versterkermarkeringen of open chromatinepieken in kransslagaderweefsel, VSMC’s of fibroblasten (aanvullende figuur 7 en aanvullende tabel 5). Van de meest geassocieerde varianten voor SCAD waren er 14 expressie-kwantitatieve eigenschaploci (eQTL’s) voor nabijgelegen genen in de aorta, kransslagader of scheenbeenslagader, volbloed of gekweekte fibroblasten (figuur 1b en aanvullende tabel 5).
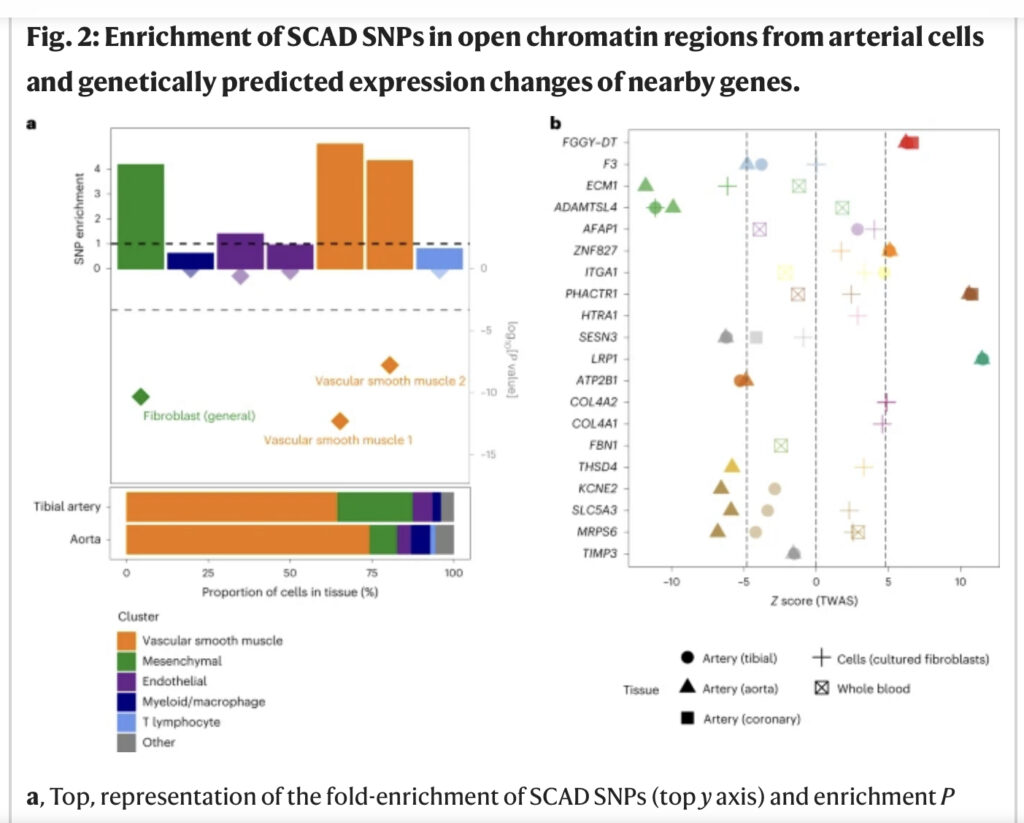
Fig. 2: Verrijking van SCAD SNP’s in open chromatinegebieden van arteriële cellen en genetisch voorspelde expressieveranderingen van nabijgelegen genen.
a, Top, weergave van de vouwverrijking van SCAD SNP’s (bovenste y-as) en verrijking P-waarde (log-schaal; onderste y-as) tussen de open chromatinegebieden van zeven eencellige subclusters die bijdragen aan> 1% van de cellen in de slagader weefsel24. De SCAD 95% geloofwaardige set van causale SNP’s en hun proxy’s voor koppelingsonevenwicht werden gematcht met willekeurige pools van naburige SNP’s met behulp van het GREGOR-pakket43. Verrijking vertegenwoordigt de verhouding van het aantal SCAD-SNP’s die open chromatinegebieden overlappen ten opzichte van het gemiddelde aantal overeenkomende SNP’s die dezelfde gebieden overlappen. P -waarden werden geëvalueerd door middel van een binominale eenzijdige test, met grotere verrijking als alternatieve hypothese43. De onderste stippellijn vertegenwoordigt de significantie (P < 0,05) na aanpassing voor 105 subclusters. Hogere ondoorzichtigheid wordt gebruikt om significante associaties te identificeren (aangepaste P < 0,05). Bodem, samenstelling van slagaderweefsels ten opzichte van 105 eencellige subclusters, zoals bepaald door snATAC-seq in 30 volwassen weefsels24. Alleen subclusters die> 1% van de cellen van de aorta of de tibiale slagader vertegenwoordigden, waren vertegenwoordigd. b, weergave van de SCAD TWAS z-score voor elk geprioriteerd gen in GWAS-loci. De puntvorm geeft het weefsel aan dat wordt gebruikt in de TWAS-associatie. De puntkleur onderscheidt genen die zich op verschillende loci bevinden. De afwezigheid van een symbool geeft aan dat het gen geen significante erfelijkheidsgraad vertoonde op basis van de eQTL-gegevens in het overeenkomstige weefsel. TWAS P-waarden werden berekend door middel van een tweezijdige z-test tegen een nulverdeling berekend door permutatie voor elk gen of weefsel44. Hogere ondoorzichtigheid wordt gebruikt om significante associaties te identificeren (Bonferroni aangepast P < 0,05), wat overeenkomt met een z score van >4,8 of <−4,8 (grijze stippellijnen).
Weefselcoagulatie als een nieuw mechanisme bij SCAD
We hebben een strategie met meerdere bronnen toegepast om kandidaatgenen te identificeren die zich bevinden in risico- of GWAS-loci, of loci die risico lopen op SCAD. We hebben prioriteit gegeven aan: (1) genen die het doelwit waren van eQTL’s die colocaliseren met een GWAS-signaal (aanvullende figuur 8a en aanvullende tabel 6) of transcriptoom-brede associatiestudie (TWAS) hits in ten minste één weefsel dat relevant is voor arteriële dissectie (aorta, coronaire of scheenbeenslagader, fibroblasten of volbloed uit de Genotype Tissue Expression (GTEx)-database) (aanvullende figuur 8b en aanvullende tabel 7); (2) genen met een biologische functie gekoppeld aan het cardiovasculaire systeem bij mensen of muizen; (3) genen die betrokken zijn bij significante chromatine-conformatie-interacties over lange afstand van Hi-C-gegevens met SCAD-geassocieerde varianten in de aorta25; en (4) die genen die het dichtst bij of overlappend zijn met de meest geassocieerde varianten. We identificeerden één specifiek en sterk kandidaat-gen in 14 loci (figuur 1b). Het weefselfactorgen F3 viel bijvoorbeeld op als het meest waarschijnlijke doelgen nabij rs1146473 (odds ratio = 1.32; P = 5.8 × 10−9) – een locus op chromosoom 1 die we beschrijven als nieuw voor SCAD en elke cardiovasculaire ziekte of eigenschap tot nu toe. F3 is het meest coderende gen voor het associatiesignaal en was een TWAS-hit in slagaderweefsel (aanvullende tabel 7). Bovendien is het rs1146473-risico-allel voor SCAD vol vertrouwen (posterieure waarschijnlijkheid = 94%) gecolokaliseerd met een eQTL-signaal van F3 in de aorta, wat het genetische risico ondersteunt om mogelijk het resultaat te zijn van verminderde F3-expressie in slagaders (figuur 2b en aanvullende tabel). 6). Weefselfactor, ook bekend als stollingsfactor III, vormt een complex met factor VIIa, de primaire initiator van bloedstolling. Daarom is verminderde factor III-expressie mogelijk een belangrijk biologisch mechanisme dat bijdraagt aan de vorming van hematoom in de kransslagaders van SCAD-overlevenden. Beschouwing van genen die coderen voor medicijnbare doelwitten, zoals afgeleid door Finan et al.26, gaf aan dat weefselfactor een geneesmiddelkandidaat in de klinische fase is (tier 1 medicijnbaar doelwit), met doelwitreferentienummers CHEMBL4081 (factor III) en CHEMBL2095194 (factor III/factor VII). complex) (aanvullende tabel 8).
Om de biologische mechanismen met geprioriteerde genen globaal te beoordelen, hebben we een netwerkquery toegepast op basis van Bayesiaanse genregulerende netwerken die zijn opgebouwd uit expressie- en genetische gegevens van arteriële weefsels en fibroblasten . We ontdekten dat extracellulaire matrixorganisatie de biologische functie is waarop de meeste geprioriteerde genen en hun respectieve directe subnetwerken geclusterd zijn (aanvullende figuur 9). Van de genen die we prioriteit hebben gegeven in nieuwe loci, codeert een aantal eiwitten die betrokken zijn bij de vorming van extracellulaire matrix, waaronder integrine alfa 1 (ITGA1), collageen type IV alfa 1-keten (COL4A1) en alfa 2-keten (COL4A2), serineprotease HtrA serinepeptidase 1 (HTRA1), metallopeptidase trombospondine type 1-domein met 4 (THSD4, coderend voor een partner van fibrilline 1, waarvan het gen zich bevindt in een eerder gerapporteerde SCAD-locus (FBN1)) en TIM metallopeptidaseremmer 3-gen (TIMP3). Interessant is dat integrine alfa 1-, HTRA1- en collageen type IV-subeenheden werden gelabeld als potentieel medicijnbare doelen op basis van hun gelijkenis met goedgekeurde medicijndoelen en leden van belangrijke geneeskrachtige genfamilies (niveau 3; aanvullende tabel 8). Merk op dat het F3-subnetwerk ook geclusterd is in extracellulaire matrixorganisatie en verbonden is met HTRA1- en TIMP3-subnetwerken via Bayesiaanse netwerkranden van de aorta en kransslagader (aanvullende figuur 9).
Gedeelde genetica tussen SCAD en arteriële aandoeningen
Met uitzondering van de F3-locus, waren SCAD-risicoloci binnen 1 megabase van de leidende SCAD-varianten op zijn minst suggestief (P < 10−5) geassocieerd met andere vormen van cardiovasculaire en neurovasculaire aandoeningen. Met behulp van kenmerkcolokalisatieanalyses ontdekten we dat dezelfde varianten waarschijnlijk causaal waren voor zowel SCAD als de andere ziekten of kenmerken op 15 loci (figuur 3a en aanvullende tabel 9). De richtingen van de effecten waren echter niet systematisch consistent over de loci voor alle ziekten.
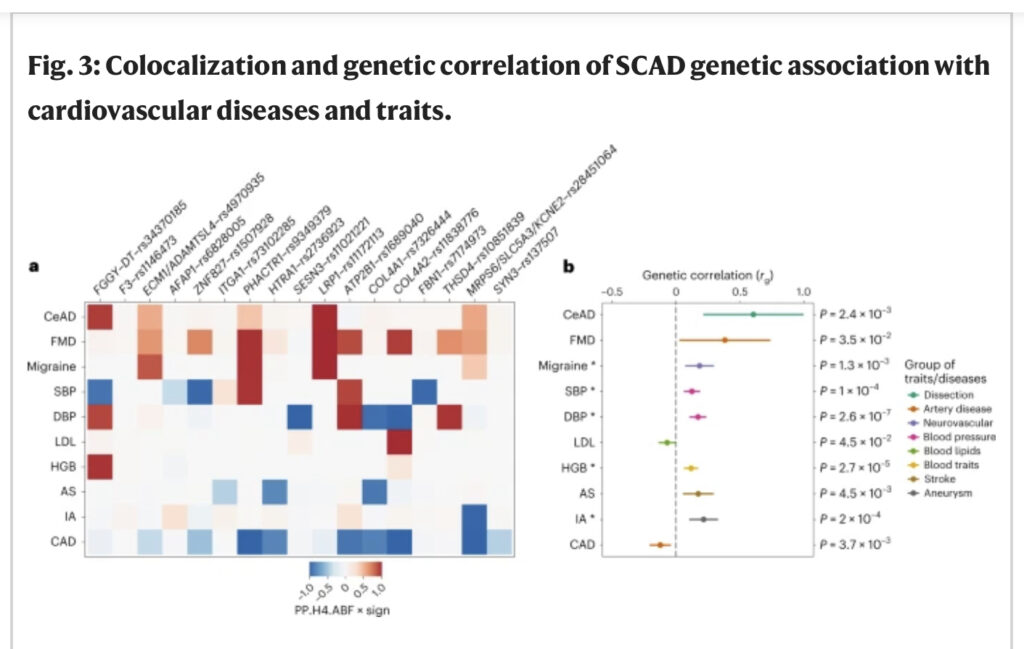
Fig. 3: Colokalisatie en genetische correlatie van SCAD-genetische associatie met hart- en vaatziekten en eigenschappen.
a, Heatmap die de colokalisatie van SCAD-signalen weergeeft met GWAS-analyse van de volgende hart- en vaatziekten of eigenschappen: cervicale arteriedissectie (CeAD), multifocale FMD, migraine, bloeddruk (SBP en DBP), LDL-cholesterolbloedconcentratie, hemoglobineconcentratie (HGB) , elke beroerte (AS), intracraniaal aneurysma (IA) en CAD. De tegelkleur vertegenwoordigt de H4-coëfficiënt van geschatte Bayes-factor (ABF) colocalisatie (dat wil zeggen, de posterieure waarschijnlijkheid van de twee kenmerken die één causale variant delen op de locus (PP.H4.ABF; 0-1)) vermenigvuldigd met het teken van colokalisatie (+1 als beide eigenschappen hetzelfde risico of een hoger gemiddeld allel hebben en -1 als het tegenovergestelde allel is)). b, Forest-plot die genetische correlaties met SCAD weergeeft. De Rho-coëfficiënt van genetische correlatie (rg), verkregen met behulp van koppelingsonevenwichtigheidsscore-regressie, wordt weergegeven op de x -as (midden van de foutbalk). Het bereik van elke balk vertegenwoordigt het 95%-BI. Ongecorrigeerde P-waarden verkregen door tweezijdige Wald-test voor genetische correlaties zijn aangegeven. Sterretjes geven significantie aan na Bonferroni-correctie voor het testen van 26 kenmerken (P < 1.9 × 10−3) (aanvullende tabel 10).
Wereldwijd toonden SCAD-loci bewijs voor een hoge posterieure waarschijnlijkheid voor dezelfde risico-allelen om waarschijnlijk ook oorzakelijk te zijn voor MKZ en halsslagaderdissectie (figuur 3a en aanvullende tabel 9). Linkage onevenwichtigheid score regressie-gebaseerde genetische correlaties gaven aan dat SCAD positief correleert met FMD (rg = 0.38 ± 0.18; P = 0.03) en halsslagader dissectie (rg = 0.61 ± 0.20; P = 2.4 × 10−3; Fig 3b en Aanvullend Tabel 10), wat consistent is met de klinische observatie van frequente coëxistentie van deze arteriopathieën bij patiënten met SCAD. FMD wordt bijvoorbeeld gemeld bij ∼ 40-60% van de patiënten met SCAD11,30. Gestratificeerde analyses in de vier grootste case-control-onderzoeken waarin FMD-arteriopathieën werden gescreend, wezen op globaal vergelijkbare associaties met SCAD (aanvullende figuur 10 en aanvullende tabel 11). Ten slotte gaven genetische correlaties aan dat SCAD positief correleert met verschillende neurovasculaire aandoeningen waarbij voornamelijk de arteriële structuur en/of functie is veranderd, waaronder beroerte (rg = 0,17 ± 0,06; P = 4,5 × 10−3), migraine (rg = 0,18 ± 0,06; P = 1,3 × 10−3), intracraniaal aneurysma (rg = 0,22 ± 0,06; P = 2,0 × 10−4) en subarachnoïdale bloeding (rg = 0,27 ± 0,07; P = 6.4 × 10−5) (figuur 3b en aanvullend Tabel 10).
Tegengestelde genetische link tussen SCAD en CAD
Terwijl patiënten met CAD overwegend mannen zijn (~75%) die vaak reeds bestaande cardiometabole comorbiditeiten hebben (voornamelijk dyslipidemie, hypertensie en diabetes type 2), zijn patiënten met SCAD gemiddeld jonger, hebben minder cardiovasculaire risicofactoren en zijn overwegend vrouw ( >90%)2,4. Met behulp van colokalisatie van genetische associatie en genetische correlatie hebben we SCAD genetisch vergeleken met CAD. We ontdekten dat van de SCAD-loci er verschillende bekend waren die verband hielden met CAD. Colokalisatieanalyses van ziekteassociaties toonden aan dat voor zes loci SCAD en CAD waarschijnlijk dezelfde causale varianten delen met hoge posterieure waarschijnlijkheden (posterieure waarschijnlijkheid van de gedeelde causale varianthypothese (H4) = 84-100%), maar allelen met tegenovergestelde risico-allelen ( Fig. 3a en aanvullende tabel 7). Genetische correlatie bevestigde een genoombrede negatieve correlatie tussen SCAD en CAD (rg = −0.12 ± 0.04; P = 3.7 × 10−3) (aanvullende tabel 10), inclusief na conditionering SCAD GWAS-resultaten op systolische bloeddruk (SBP) of diastolische Bloeddruk (DBP) GWAS-resultaten met behulp van de multitrait-gebaseerde voorwaardelijke en gewrichtsanalyse (MTCOJO) Tool31 (RGCAD/SBP = −0.19 ± 0,04 (P = 4,6 × 10−6); RGCAD/DBP = −0.19 ± 0,04 (P = 1.3 × 10−5)) (aanvullende tabel 12 en aanvullende afbeelding 11).
Cardiovasculaire risicofactoren en risico op SCAD en CAD
We ontdekten dat SCAD verschillende causale varianten deelde met SBP en DBP, waarbij zowel dezelfde als tegengestelde directionele effecten betrokken waren (figuur 3a en aanvullende tabel 9). We vonden één gedeelde locus met hemoglobinewaarden en een significante genetische correlatie met SCAD (rg = 0.12 ± 0.03; P = 2.7 × 10−5; Fig. 3b). SCAD-loci werden echter niet gedeeld met body mass index (BMI), lipidenkenmerken (waaronder low-density lipoprotein (LDL) cholesterol en high-density lipoprotein (HDL)), diabetes type 2 of roken, en deze kenmerken correleerden niet met SCAD op genomisch niveau (aanvullende tabellen 9 en 10). Interessant is dat we significante positieve genetische correlaties vonden met zowel SBP (rg = 0.12 ± 0.03; P = 1.0 × 10−4) als DBP (rg = 0.17 ± 0.03; P = 2.6 × 10−7), wat wijst op een gedeelde genetische basis met SCAD (figuur 3b en aanvullende tabel 10). Om te beoordelen in hoeverre bloeddruk en de belangrijkste cardiovasculaire risicofactoren kunnen bijdragen aan het risico op SCAD, hebben we bestaande GWAS-datasets gebruikt om instrumentele variabelen te identificeren en vergelijkende Mendeliaanse randomisatie-associaties met SCAD of CAD uitgevoerd. We vonden robuuste significante associaties geschat door inverse variantie-gewogen (IVW), MR-Egger en gewogen mediaanmethoden tussen genetisch voorspelde bloeddrukkenmerken en verhoogd risico op SCAD (βIVW/SBP = 0.05 ± 0.01 (P = 7.6 × 10−6) βIVW/DBP = 0,10 ± 0,02 (P = 1,9 × 10−8)) en CAD (βIVW/SBP = 0,04 ± 0,002 (P = 8,6 × 10−49); BP = 0.06 ± 0.004 (P = 1.6 × 10-44)) (Afb. 4 en aanvullende tabel 13). Vergelijkbare associaties werden geschat toen we alleen vrouwen met SCAD, vrouwen met CAD of mannen met CAD analyseerden, hoewel analyses alleen bij mannen met SCAD beperkt waren door het extreem kleine aantal mannelijke gevallen (aanvullende tabel 14). Genetisch bepaalde BMI, lipidenkenmerken, diabetes type 2 en rookstatus hadden geen invloed op het risico op SCAD. We konden echter bevestigen dat deze cardiometabole eigenschappen sterke genetische risicofactoren zijn voor CAD (figuur 4 en aanvullende tabel 13). Onze bevindingen geven aan dat genetisch verhoogde bloeddruk de enige gedeelde genetische risicofactor is tussen SCAD en CAD, zij het met potentieel verschillende genetische loci.
Fig. 4: Mendeliaanse randomisatie-associaties tussen de belangrijkste cardiovasculaire risicofactoren en SCAD of CAD.
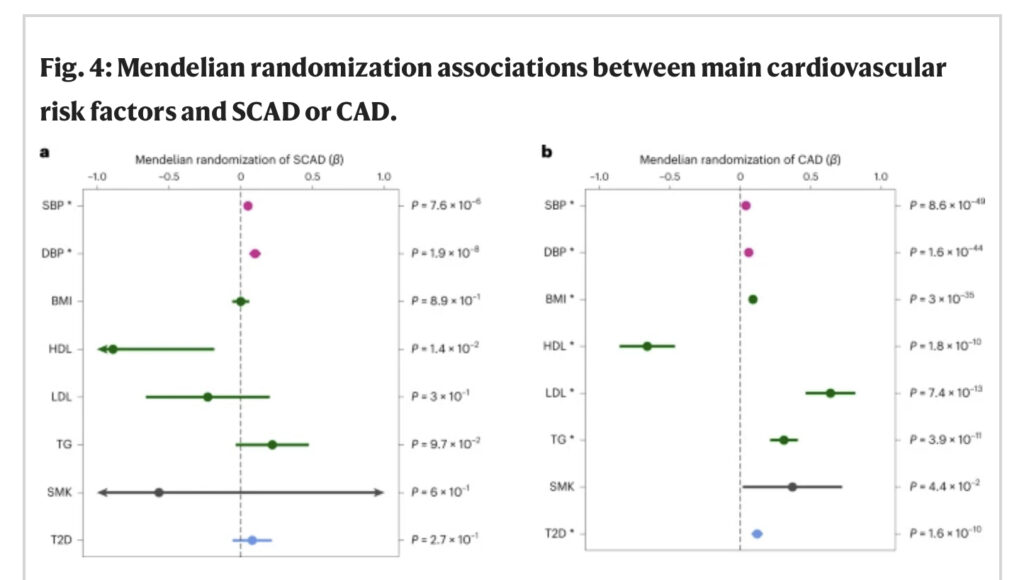
a, b, Forest-plots die Mendeliaanse randomisatie-associaties tussen cardiovasculaire risicofactoren en SCAD (ncases = 1.917; ncontrols = 9.292) (a) of CAD (ncases = 181.522; ncontrols = 984.168) (b) weergeven. Associatieschattingen (β; midden van de foutbalken) verkregen uit Mendeliaanse randomisatieanalyses met behulp van de IVW-methode worden weergegeven op de x -as. Het bereik van elke balk vertegenwoordigt het 95%-BI. Ongecorrigeerde P -waarden van de associaties verkregen door tweezijdige Wald-test worden aangegeven. n = 340.159 (SBP), 340.162 (DBP), 359.983 (BMI), 315.133 (HDL), 343.621 (LDL), 343.992 (triglyceriden (TG)), 164.638 gevallen en 195.068 controles (roken (SMK)) en 74.124 gevallen en 824.006 controles (diabetes type 2 (T2D)). De sterretjes geven significantie aan na Bonferroni-correctie voor het testen van negen eigenschappen (P < 5.6 × 10−3) (aanvullende tabel 13).
Discussie
In dit artikel geven we de grootste studie tot nu toe gericht op het begrijpen van de genetische basis van SCAD – een onderbelichte oorzaak van AMI die voornamelijk vrouwen treft. We rapporteren nieuwe associaties en demonstreren een hoge polygene erfelijkheidsgraad voor SCAD. We maken gebruik van integratieve functionele annotaties om prioriteit te geven aan genen die waarschijnlijk worden gereguleerd in VSMC’s en de fibroblasten van slagaders. Inzichten uit de biologische functies van genen benadrukken de centrale rol van extracellulaire matrixintegriteit en onthullen verminderde weefselcoagulatie als een nieuw potentieel mechanisme voor SCAD. Wereldwijd tonen we aan dat de polygene basis van SCAD wordt gedeeld met een belangrijke reeks hart- en vaatziekten. Er wordt echter een opvallend tegengestelde genetische impact gevonden bij atherosclerotische CAD, waarbij meerdere risicoloci betrokken zijn en die leidt tot een genoombrede negatieve genetische correlatie. We leveren bewijs ter ondersteuning van genetisch voorspelde hogere bloeddruk als een belangrijke risicofactor voor SCAD, maar niet voor andere gevestigde cardiovasculaire factoren. Onze resultaten vormen de basis voor toekomstig onderzoek naar nieuwe biologische routes die relevant zijn voor zowel SCAD als CAD en potentiële therapeutische en preventieve strategieën die specifiek gericht zijn op SCAD.
Als een onderbelichte aandoening waarvan eerder werd gedacht dat het ongebruikelijk was, werd aanvankelijk vermoed dat SCAD zeldzame en zeer penetrante mutaties met zich meebracht. Recente sequencing-onderzoeken hebben echter gesuggereerd dat slechts een klein deel (~ 3,5%) van de SCAD-gevallen te wijten is aan zeldzame varianten16,32. Dit is in overeenstemming met de toenemende klinische erkenning die suggereert dat deze aandoening niet zeldzaam is en wereldwijd voorkomt in populaties van zowel Europese als niet-Europese afkomst, met vergelijkbare ziektekenmerken en waarschijnlijk vergelijkbare prevalentie2,4,33,34. Ondanks een bescheiden steekproefomvang, hebben we 16 risicoloci geïdentificeerd die verantwoordelijk zijn voor ongeveer een kwart van de polygene erfelijkheid, die naar onze schatting zo hoog is als ~ 71%, wat aangeeft dat SCAD overwegend een complexe polygene ziekte is. We erkennen echter dat grotere GWAS-instellingen, waaronder voorouderlijk diverse populaties, de statistische kracht zullen vergroten die nodig is om validatie te bieden door replicatie van de gerapporteerde risico-loci en geschatte polygene erfelijkheid.
Deze studie ondersteunt de aanwezigheid van genetische overlap tussen het risico op SCAD en andere vasculaire ziekten waarbij over het algemeen jongere individuen en meer vrouwen betrokken zijn, zoals cervicale arteriële dissectie, migraine, subarachnoïdale bloeding en FMD. Van deze aandoeningen is gemeld dat ze vaker voorkomen bij patiënten met SCAD10,11,12,13, wat gedeelde causale biologische mechanismen ondersteunt. Van de genen die we prioriteren als nieuwe SCAD-loci, benadrukken we het ATPase-plasmamembraan Ca2+ dat 1 gen transporteert (ATP2B1) waarvan we onlangs hebben gemeld dat het associeert met FMD35 – een gevestigde locus voor bloeddrukrisico36 via zijn rol in intracellulaire calciumhomeostase in VSMC’s en bloeddrukregulatie37. Het belangrijkste is dat we bewijs leveren voor een oorzakelijk genetisch effect van zowel SBP als DBP op SCAD-risico. Deze bevindingen vormen een belangrijke genetische basis ter ondersteuning van observationele gegevens die suggereren dat controle van de bloeddruk een belangrijke factor kan zijn bij het verminderen van het risico op herhaling na SCAD38. Onze bevindingen suggereren echter ook dat het beheersen van andere oorzakelijke risicofactoren voor CAD, zoals LDL-cholesterol met statines, mogelijk minder voordeel oplevert bij SCAD dan bij CAD.
Kennis van de moleculaire mechanismen die leiden tot SCAD is beperkt. Inzichten uit sequentiestudies van zeldzame genetische varianten hebben aangetoond dat de meeste geassocieerd zijn met genen die bekend zijn van erfelijke bindweefselaandoeningen zoals vasculaire Ehlers-Danlos-, Loeys-Dietz- en Marfan-syndromen, evenals polycysteuze nierziekte bij volwassenen16,32. Een opvallende bevinding uit ons onderzoek is de identificatie van het weefselfactorgen F3 – een cruciaal onderdeel van weefselgemedieerde bloedstolling – als een sterk kandidaat-gen in een risicolocus voor SCAD. We ontdekten dat genetisch bepaalde lagere expressie van F3 in arterieel weefsel geassocieerd was met een hoger risico op SCAD, waarbij varianten betrokken waren die zich bevinden in vermeende functionele regulerende elementen in de kransslagader, VSMC’s en fibroblasten. Weefselfactor wordt gesynthetiseerd op het subendotheliale niveau van VSMC’s en door fibroblasten in de adventitia die de slagaders omringen39. Wanneer bij SCAD een intramurale bloeding eenmaal is begonnen, kunnen voortplanting en drukverhoging van het valse lumen gedeeltelijk afhangen van coagulatie en stabilisatie van het hematoom. Weefselfactor is ook een medicijnbaar doelwit, zij het een potentieel uitdagend doelwit, gezien de bekende meerdere fysiologische en pathofysiologische rollen, variërend van hemostase tot metastase van kanker. Weefselfactor wordt uitgebreid bestudeerd in de context van protrombotische aandoeningen, waaronder atherosclerose, hoewel met name de genetische varianten die we hier beschrijven niet geassocieerd zijn met atherosclerotische ziekte. Dit kenmerk is een uitzondering op de zeer pleiotrope aard van de varianten die we beschrijven in de resterende SCAD-loci, wat duidt op verminderde weefsel-geïnitieerde coagulatie als een vermeend specifiek mechanisme in SCAD.
We identificeren regulatie van de extracellulaire matrix van slagaders als het overheersende polygene biologische mechanisme voor SCAD. Integratieve prioriteringsanalyses onthulden 13 potentiële oorzakelijke genen met gevestigde sleutelrollen bij het handhaven van de integriteit en functie van de arteriële wand. Hiervan belichten we de serineprotease HTRA1 en metallopeptidaseremmer TIMP3, die betrokken zijn bij de demontage van de matrix. TIMP3 clustert in het hoofdnetwerk voor extracellulaire matrixorganisatie die ADAMTSL4, LRP1 en COL4A1 omvat, met verbindingen met subnetwerken van F3. Deze clustering is consistent met de biologische functie van TIMP3 als een remmer van matrixmetalloproteïnasen met domeinen die interageren met ADAMTS-eiwitten en LRP1, waarbij eiwitten zijn betrokken die worden gecodeerd door genen die prioriteit hebben in SCAD-loci40. Interessant is dat we een nieuw associatiesignaal met SCAD hebben gevonden in het metallopeptidase trombospondine type 1-domein met 4-gen (THSD4) dat fibrilline 1 elastische vezelassemblage bevordert, en bevestigen de eerder gerapporteerde associaties nabij ADAMTSL4 en FBN1 (refs. 18, 20). We toonden aan dat genetisch verlaagde expressies van deze genen in slagaders gecorreleerd waren met allelen met een hoger SCAD-risico in slagaders of fibroblasten. Deze bevinding suggereert dat een genetische aanleg voor een zwakkere extracellulaire matrix de kwetsbaarheid van het passeren van intramurale microvaatjes voor verstoring kan vergroten, waardoor het risico op initiatie en verspreiding van een vals lumen binnen de coronaire vaatwand toeneemt, wat leidt tot SCAD.
Veel van de risico-loci voor SCAD die we hier rapporteren, evenals hun geprioriteerde genen, zijn al bekend van GWAS’s voor atherosclerotische ziekten. Hier leveren we echter overtuigend en intrigerend bewijs voor de tegenovergestelde directionaliteit van een substantiële fractie van genetische bases voor SCAD versus CAD, wat suggereert dat sommige belangrijke biologische mechanismen die betrokken zijn bij de twee ziekten waarschijnlijk ook tegengesteld zijn, wat consistent is met de klinische observatie van een lager dan verwachte last van atherosclerotische ziekte bij patiënten met SCAD. De associatiesignalen in de COL4A1/COL4A2-locus zijn bijvoorbeeld in de tegenovergestelde richting van hun bijdrage aan CAD41. Deze locus codeert voor α1- en α2-ketens van type IV collageen, met transcripten gegenereerd door een gemeenschappelijke promotor. Type IV collageen is het hoofdbestanddeel van het basismembraan van arteriële cellen en speelt een sleutelrol in de structurele integriteit en biologische functies van VSMC’s in de tunica muscularis. Verminderde expressie van collageen IV verhoogt het risico op CAD15,42. Voorgestelde mogelijke mechanismen hiervoor zijn onder meer een ontremming van VSMC-intimale migratie tijdens atherogenese of een toename van de kwetsbaarheid van atherosclerotische plaque voor scheuren42. In tegenstelling tot CAD geven onze gegevens aan dat genetisch gemedieerde verhoogde expressie van collageen IV ook het risico op SCAD verhoogt. Een beter begrip van hoe deze tegengestelde richtingsveranderingen het risico op CAD en SCAD wijzigen, heeft een aanzienlijk potentieel om ons begrip van de moleculaire genetische mechanismen die risico’s met zich meebrengen bij beide ziekten te vergroten.
methoden
Patiënten en controlepopulaties
Onze meta-analyse omvatte deelnemers van Europese afkomst uit acht studies: DISCO-3C, SCAD-UK I, SCAD-UK II, Mayo Clinic, DEFINE-SCAD, CanSCAD/MGI, VCCRI I en VCCRI II (aanvullende figuur 1). Patiënten met SCAD vertoonden vergelijkbare klinische kenmerken (aanvullende tabel 1), evenals homogene diagnose-, uitsluitings- en inclusiecriteria. Alle onderzoeken zijn goedgekeurd door nationale en/of institutionele ethische beoordelingsraden. Verdere studiespecifieke klinische details zijn te vinden in de aanvullende opmerking.
Genoombrede associatie-meta-analyse
Details van de pre-imputatie kwaliteitscontrolestappen voor elk onderzoek zijn opgesomd in aanvullende tabel 15. In het kort werd genotypering uitgevoerd met behulp van in de handel verkrijgbare arrays of genoomsequencing (SCAD-UK II en VCCRI II). Om het aantal geteste SNP’s en de overlapping van varianten die beschikbaar zijn voor analyse tussen verschillende arrays te vergroten, werden de genotypen van alle cohorten van Europese voorouders, behalve SCAD-UK II en VCCRI II, toegerekend aan het Haplotype Reference Consortium versie 1.1 referentiepanel45 op de Michigan Imputation Server46 . In elk onderzoek werd een GWAS uitgevoerd onder een additief genetisch model met behulp van PLINK versie 2.0 (ref. 47). Voor chromosoom X bevonden mannen en vrouwen zich beide op een schaal van 0,2 volgens het chromosoom X-inactiveringsaannamemodel. Modellen werden aangepast voor populatiestructuur met behulp van residuen van de eerste vijf hoofdcomponenten en geslacht, behalve in de analyses voor alleen vrouwen. Vóór de meta-analyse hebben we SNP’s verwijderd met lage kleine allelfrequenties (<0,01), lage imputatiekwaliteit (r2 < 0,8) en afwijkingen van het Hardy-Weinberg-evenwicht (P < 10−5). In totaal voldeden 6.691.677 varianten aan deze criteria en werden behouden in de einduitslag.
Resultaten van individuele GWAS’s werden gecombineerd met behulp van een inverse variantie-gewogen meta-analyse met vaste effecten in METAL-software48, met correctie voor genomische controle. Heterogeniteit werd beoordeeld met behulp van de I2-metriek uit de volledige meta-analyse op studieniveau. Heterogeniteit tussen studies werd getest met behulp van de Q-statistiek van Cochran en werd als significant beschouwd bij P ≤ 10−3. De genoombrede significantiedrempel was vastgesteld op het niveau van P = 5.0 × 10−8. LocusZoom (//locuszoom.org/) werd gebruikt om de resultaten regionaal te visualiseren.
Functionele annotatie
Identificatie van mogelijke functionele varianten
Om een lijst met mogelijke functionele varianten te genereren, hebben we eerst de 95% geloofwaardige reeks varianten geïdentificeerd met behulp van de ppfunc-functie van het corrcoverage R-pakket (versie 1.2.1). De posterieure waarschijnlijkheid van causaliteit werd geëvalueerd op basis van marginale z scores voor alle varianten binnen 500 kilobases (kb) van de leidende SNP op elke locus. In de COL4A1/COL4A2-locus, waar we twee associatiesignalen vonden, werden deze gescheiden door een grens op gelijke afstand van elke lead-SNP te plaatsen voor de opname van SNP’s in de analyse. Varianten met een gecumuleerde posterieure waarschijnlijkheid tot 95% werden behouden voor verdere analyse. Om potentieel slecht toegerekende varianten in een van de individuele case-control-onderzoeken te overwegen, hebben we ook varianten opgenomen in onevenwichtigheid met hoge koppeling (r2 > 0.7) met de leidende SNP op elke locus, op basis van informatie van Europese populaties (1000 Genomes-referentiepanel) opgevraagd gebruik van de ldproxy-functie van het LDlinkR-pakket (versie 1.1.2)49.
Verrijking van SCAD-varianten in regelgevende regio’s
Om de verrijking van SCAD-geassocieerde SNP’s tussen functioneel geannoteerde genomische regio’s te berekenen, hebben we beschikbare H3K27ac-chromatine-immunoprecipitatie opgehaald gevolgd door sequencing (ChIP-seq) datasets (narrowPeak-bedden) in elk weefsel van ENCODE (//www.encodeproject.org/ (ref. 50)) en single-nucleus assay voor transposase-toegankelijk chromatine met sequencing (snATAC-seq) piekbestanden (bedformaat) van de Human Enhancer Atlas (//catlas.org/humanenhancer (ref. 24)) . Een volledige lijst met gegevenssets is beschikbaar in aanvullende tabel 16. Voor H3K27ac-markeringen werden bedbestanden die overeenkomen met hetzelfde weefsel samengevoegd en gesorteerd voordat overlappende pieken werden gecombineerd met behulp van de samenvoegopdracht bedtools (versie 2.29.0). Variantverrijking werd berekend met behulp van het GREGOR-pakket (versie 1.4.0)43. Alle potentiële functionele varianten (95% geloofwaardige proxy’s voor set- en koppelingsonevenwichtigheid zoals hierboven beschreven) werden gebruikt als invoer en de parameters werden aangepast om geen aanvullende proxy’s voor koppelingsonevenwichtigheid te kiezen (LDWINDOWSIZE = 1). P -waarden werden aangepast voor meervoudige testen door Bonferroni-correctie toe te passen.
Identificatie van varianten met potentiële regulerende functie
We gebruikten H3K27ac-pieken in kransslagaders (zoals hierboven beschreven), open chromatinegebieden in gezonde kransslagaders (verkregen zoals eerder beschreven35,51) en open chromatinegebieden van samengevoegde snATAC-seq-clusters, die in kaart gebrachte fragmenten waren van snATAC-seq in 25 volwassen weefsels die we hebben opgehaald uit de Gene Expression Omnibus (GSE184462)24 in bedformaat. In kaart gebrachte fragmenten van alle clusters die> 1% van de cellen vertegenwoordigen in ten minste één arterieel weefsel (T-lymfocyt 1, CD8 +, endotheliaal algemeen 2, endotheel algemeen 1, macrofaag algemeen, fibroblast algemeen, vasculaire gladde spier 2 of vasculaire gladde spier 1) werden geëxtraheerd en gegroepeerd op geannoteerd celtype als respectievelijk T-lymfocyten, macrofagen, fibroblasten, endotheelcellen en VSMC’s. Genoomdekking werd berekend met behulp van de dekkingsfunctie van bedtools (versie 2.29.0). We hebben pieken van bedGraph-uitvoer gedetecteerd met behulp van de MACS2 bdgpeakcall-functie (Galaxy-versie 2.1.1.20160309.0) op de Galaxy-webserver52,53. Alle piekbestanden werden 100 basenparen stroomopwaarts en stroomafwaarts verlengd met behulp van de slop-functie van bedtools (versie 2.29.0). We hebben overlappingen van SCAD-potentiële functionele varianten met relevante genomische regio’s gedetecteerd met behulp van de functie findOverlap uit het rtracklayer-pakket (versie 1.52.1)54. We gebruikten de Integrated Genome Browser (versie 9.1.8) om leesdichtheidsprofielen en piekposities te visualiseren in de context van het menselijk genoom55.
Prioritering van genen
Genen die zich binnen 500 kb van leadvarianten bevonden, werden geannoteerd om prioriteit te geven aan de meest waarschijnlijke causale genen. Om de dichtstbijzijnde genen te vinden van leidende SNP’s en genen die overlappen met varianten in de geloofwaardige set van causale SNP’s, werden gencoördinaten opgehaald uit Gencode release 38 en uitgelijnd met hg19 genomische coördinaten (gencode.v38lift37.annotation.gff3.gz). Significante eQTL-associaties en alle SNP-gene eQTL-associaties in versie 8 van de GTEx-database zijn opgehaald van de GTEx-website (www.gtexportal.org/home/datasets). Colocalisatie van associatie met SCAD en eQTL’s werd geëvalueerd met behulp van het R coloc-pakket (versie 5.1.0) met standaardwaarden als priors. We waren van mening dat er bewijs was voor colokalisatie als H4-coëfficiënten> 75% waren of als eQTL-associatie significant was voor SCAD-lead-SNP’s en H4 meer dan 25% was. TWAS’s werden uitgevoerd met behulp van het FUSION R/Python-pakket44. Genexpressiemodellen werden vooraf berekend op basis van GTEx-gegevens (versie 8 release) en werden geleverd door de auteurs. Alleen genen met een erfelijkheidsgraad P < 0,01 werden in de analyse gebruikt. Beide tools maakten gebruik van koppelingsonevenwichtsinformatie van het Europese panel van fase 3 van het 1000 Genomes Project. Bonferroni meervoudige testcorrectie werd toegepast met behulp van de p.adjust-functie in R (versie 4.1.0). Significante capture Hi-C-hits in aortaweefsel werden verstrekt als aanvullende gegevens door Jung et al.25. Genen geassocieerd met cardiovasculaire fenotypes van muizen (code MP:0005385) werden opgehaald uit de Mouse Genome Informatics-database (www.informatics.jax.org)56. We hebben ook de DisGeNET-database doorzocht, met behulp van het disgenet2r-pakket (versie 0.99.2), voor genen met gerapporteerd bewijs in hart- en vaatziekten bij de mens (code C14) met een score van> 0,2, inclusief “ALLE” databases57. Bij afwezigheid van een missense-variant kregen colokalisatie- en TWAS-criteria een tienvoudig gewicht in vergelijking met andere criteria. Op elke locus hebben we prioriteit gegeven aan genen die aan het grootste aantal criteria voldoen. In gevallen waarin meerdere kandidaten werden behouden, hebben we prioriteit gegeven aan genen die het meest waarschijnlijk een functie hebben bij arteriële ziekte (bijvoorbeeld expressie in arteriële weefsels of uitsluiting van pseudo-genen).
Druggabiliteit van geprioriteerde genen
De medicijnbaarheid van de genproducten die via de GWAS zijn geïdentificeerd, werd beoordeeld door te verwijzen naar de set genen die coderen voor medicijnbare doelen afgeleid door Finan et al.26 met behulp van ChEMBL versie 17. Doelen in deze set zijn onderverdeeld in: (1) de werkzaamheidsdoelen van goedgekeurde middelen en kandidaat-geneesmiddelen in de klinische fase (tier 1); (2) genen die coderen voor doelwitten met bekende bioactieve geneesmiddelachtige bindingspartners van kleine moleculen en genen met een substantiële sequentie met goedgekeurde geneesmiddeldoelwitten (tier 2); en (3) genen die coderen voor uitgescheiden of extracellulaire eiwitten, eiwitten met een verder verwijderde gelijkenis met goedgekeurde medicijndoelen en leden van belangrijke geneeskrachtige genfamilies die nog niet zijn opgenomen in niveaus 1 of 2. Verdere opzoekingen van goedgekeurde en klinische fasedoelen werden uitgevoerd tegen ChEMBL58 versie 30 en het British National Formulary (geraadpleegd op 9 april 2021). Merk op dat geïdentificeerde medicijndoelwitten kunnen zijn: (1) een enkel eiwit dat een 1:1-link verschaft met het oorzakelijke gen dat is genomineerd in een GWAS- en post-GWAS-analyse; (2) een eiwitcomplex waarbij het causale gen kan coderen voor een lid van het complex; of (3) een eiwitfamilie waarvan het oorzakelijke gen een lid van de familie is.
Prioritering van genen
Genen die zich binnen 500 kb van leadvarianten bevonden, werden geannoteerd om prioriteit te geven aan de meest waarschijnlijke causale genen. Om de dichtstbijzijnde genen te vinden van leidende SNP’s en genen die overlappen met varianten in de geloofwaardige set van causale SNP’s, werden gencoördinaten opgehaald uit Gencode release 38 en uitgelijnd met hg19 genomische coördinaten (gencode.v38lift37.annotation.gff3.gz). Significante eQTL-associaties en alle SNP-gene eQTL-associaties in versie 8 van de GTEx-database zijn opgehaald van de GTEx-website (www.gtexportal.org/home/datasets). Colocalisatie van associatie met SCAD en eQTL’s werd geëvalueerd met behulp van het R coloc-pakket (versie 5.1.0) met standaardwaarden als priors. We waren van mening dat er bewijs was voor colokalisatie als H4-coëfficiënten> 75% waren of als eQTL-associatie significant was voor SCAD-lead-SNP’s en H4 meer dan 25% was. TWAS’s werden uitgevoerd met behulp van het FUSION R/Python-pakket44. Genexpressiemodellen werden vooraf berekend op basis van GTEx-gegevens (versie 8 release) en werden geleverd door de auteurs. Alleen genen met een erfelijkheidsgraad P < 0,01 werden in de analyse gebruikt. Beide tools maakten gebruik van koppelingsonevenwichtsinformatie van het Europese panel van fase 3 van het 1000 Genomes Project. Bonferroni meervoudige testcorrectie werd toegepast met behulp van de p.adjust-functie in R (versie 4.1.0). Significante capture Hi-C-hits in aortaweefsel werden verstrekt als aanvullende gegevens door Jung et al.25. Genen geassocieerd met cardiovasculaire fenotypes van muizen (code MP:0005385) werden opgehaald uit de Mouse Genome Informatics-database (www.informatics.jax.org)56. We hebben ook de DisGeNET-database doorzocht, met behulp van het disgenet2r-pakket (versie 0.99.2), voor genen met gerapporteerd bewijs in hart- en vaatziekten bij de mens (code C14) met een score van> 0,2, inclusief “ALLE” databases57. Bij afwezigheid van een missense-variant kregen colokalisatie- en TWAS-criteria een tienvoudig gewicht in vergelijking met andere criteria. Op elke locus hebben we prioriteit gegeven aan genen die aan het grootste aantal criteria voldoen. In gevallen waarin meerdere kandidaten werden behouden, hebben we prioriteit gegeven aan genen die het meest waarschijnlijk een functie hebben bij arteriële ziekte (bijvoorbeeld expressie in arteriële weefsels of uitsluiting van pseudo-genen).
Druggabiliteit van geprioriteerde genen
De medicijnbaarheid van de genproducten die via de GWAS zijn geïdentificeerd, werd beoordeeld door te verwijzen naar de set genen die coderen voor medicijnbare doelen afgeleid door Finan et al.26 met behulp van ChEMBL versie 17. Doelen in deze set zijn onderverdeeld in: (1) de werkzaamheidsdoelen van goedgekeurde middelen en kandidaat-geneesmiddelen in de klinische fase (tier 1); (2) genen die coderen voor doelwitten met bekende bioactieve geneesmiddelachtige bindingspartners van kleine moleculen en genen met een substantiële sequentie met goedgekeurde geneesmiddeldoelwitten (tier 2); en (3) genen die coderen voor uitgescheiden of extracellulaire eiwitten, eiwitten met een verder verwijderde gelijkenis met goedgekeurde medicijndoelen en leden van belangrijke geneeskrachtige genfamilies die nog niet zijn opgenomen in niveaus 1 of 2. Verdere opzoekingen van goedgekeurde en klinische fasedoelen werden uitgevoerd tegen ChEMBL58 versie 30 en het British National Formulary (geraadpleegd op 9 april 2021). Merk op dat geïdentificeerde medicijndoelwitten kunnen zijn: (1) een enkel eiwit dat een 1:1-link verschaft met het oorzakelijke gen dat is genomineerd in een GWAS- en post-GWAS-analyse; (2) een eiwitcomplex waarbij het causale gen kan coderen voor een lid van het complex; of (3) een eiwitfamilie waarvan het oorzakelijke gen een lid van de familie is.
Mendeliaanse randomisatieanalyses
We hebben een streng selectieproces toegepast voor instrumentele variabelen om de validiteit van onze Mendeliaanse randomisatieresultaten te waarborgen. Om geldige instrumentele variabelen te selecteren die de drie belangrijkste aannames respecteren ((1) sterke associatie met de blootstelling; (2) onafhankelijkheid van potentiële confounders tussen de blootstelling en uitkomst; en (3) invloed op de uitkomst alleen door de blootstelling), gebruikten we koppelingsonevenwicht klonteren met een P waardedrempel van <5 × 10−8 en een koppelingsonevenwichtigheid r2 < 0,001 binnen een venster van 10.000 kb op basis van de Europese bevolking in het 1000 Genomes Project. We hebben kandidaat-instrumentele variabelen uitgesloten die afwezig waren in de samenvattende statistische gegevens van een GWAS van onze uitkomst (SCAD/CAD). Om het risico op horizontale pleiotropie te minimaliseren, hebben we kandidaat-instrumentele variabelen verwijderd die geassocieerd waren met de uitkomst of met een hoge tot matige koppelingsonevenwichtigheid (r2 > 0,6 binnen een venster van 10.000 kb).
We gebruikten de multiplicatieve random-effects IVW-methode59 geïmplementeerd in het TwoSampleMR R-pakket om de associaties te schatten van genetisch voorspelde cardiovasculaire risicofactoren, waaronder bloeddruk (SBP en DBP), lipiden (HDL, LDL en triglyceriden), BMI, aansprakelijkheid voor roken en type 2 diabetes, met elk van de uitkomsten van belang (SCAD of CAD). Schattingen werden geschaald naar een verdubbeling van het genetisch voorspelde risico op roken, of naar een toename van één eenheid in het genetisch voorspelde kenmerk voor de continue kenmerken. We hebben gevoeligheidsanalyses uitgevoerd met behulp van de gewogen mediaan- en MR-Egger-methoden om de consistentie van schattingen te beoordelen onder alternatieve aannames over genetische pleiotropie, zoals aanbevolen59. We hebben ook de Q-test van Cochran uitgevoerd om de heterogeniteit tussen schattingen verkregen met verschillende varianten te beoordelen. Aangezien 11 risicofactoren werden beoordeeld, werd een Bonferroni-gecorrigeerd significantieniveau van 0,05/9 = 5,6 × 10−3 gebruikt als de drempel voor statistische significantie in deze analyse. P -waarden tussen 5,6 × 10−3 en 0,05 werden als suggestief significant beschouwd.
.

Geef een reactie